Il y a, dans le cerveau humain d’aujourd’hui, certaines structures nerveuses qui sont apparues il y a très longtemps, d’autres plus tard, et d’autres encore plus récemment. Dès les années 1960, le neuroanatomiste Paul MacLean avait popularisé cette approche évolutive avec son cerveau à trois étages, le reptilien, le limbique et le néocortex. J’ai écrit ailleurs pourquoi on sait aujourd’hui qu’il s’agit d’une simplification dépassée. Mais comme souvent, ce premier débroussaillage allait être ensuite raffiné par des gens comme Jaak Panksepp dont j’ai aussi déjà parlé des considérations évolutives dans la genèse des émotions qui font la part belle aux structures sous-corticales communes à tous les mammifères. Aujourd’hui on va s’intéresser à un chercheur contemporain qui approfondit encore davantage cette tradition. Il s’agit de Paul Cisek que j’ai eu la chance de voir quelques fois en conférence puisqu’il travaille à l’université de Montréal. J’ai donc déjà parlé de ses travaux dans ce blogue ici, là et encore là. Plus récemment, en 2019, Cisek a publié un articles intitulé Resynthesizing behavior through phylogenetic refinement qui poursuit la réflexion de ses travaux antérieurs sur l’origine phylogénétique de nos comportements. Je vous propose cette semaine un survol de cet article qui m’a grandement intéressé parce qu’il m’a rappelé un certain cours durant mon baccalauréat en biologie sur la morphologie évolutive des vertébrés. Cours que j’avais adoré mais trouvé très difficile. Vous me pardonnerez donc, je l’espère, ce retour aux sources un peu nostalgique. Je vais essayer de vous rendre cela le plus digeste possible. Et comme c’est dense et assez long, on terminera la semaine prochaine.
À la base de la démarche de Cisek, il y a l’idée que les catégories conceptuelles qu’on utilise depuis toujours pour classer et pour étudier les comportements ne sont peut-être pas les meilleures quand vient le temps d’essayer d’en décortiquer les mécanismes sous-jacents dans le cerveau. C’est que ces concepts sont souvent issus de l’histoire de la psychologie, à une époque où l’on ne pouvait pas vraiment regarder dans le cerveau avec les outils des neurosciences. Avec l’accessibilité de l’imagerie cérébrale dans les années 1990, on s’est rendu compte qu’il était très difficile, voire impossible, de trouver des régions ou des circuits nerveux bien définis associés par exemple à la moindre émotion. Même chose pour des concepts couramment utilisés en sciences cognitives comme l’attention, la mémoire de travail ou la prise de décision. Il y a beaucoup de débats sur les réseaux cérébraux qui les sous-tendent parce que ce n’est pas évident de leur associer des structures cérébrales particulières.
Ou alors prenez le concept même de cognition, qui est ce que tout bon neurobiologiste va avoir tendance à placer spontanément entre la perception et l’action. Est-ce que c’est vraiment comme ça que les cerveaux sont structurés ? Ou est-ce qu’on ne pourrait pas développer une taxonomie différentes qui reflèterait davantage les mécanismes biologiques qui se sont mis en place progressivement ? Et qui auraient donc une meilleure adéquation avec ces circuits. Poser la question, c’est un peu y répondre, comme on dit. Mais ça dit pas comment on pourrait arriver à les redéfinir, ces nouveaux concepts. D’où la récapitulation généalogique vraiment très instructive que propose l’article de Cisek. Mais pas juste des générations qui nous ont précédées : de tous nos ancêtres animaux !
Abonnez-vous à notre infolettre!
Pour ne rien rater de l'actualité scientifique et tout savoir sur nos efforts pour lutter contre les fausses nouvelles et la désinformation!
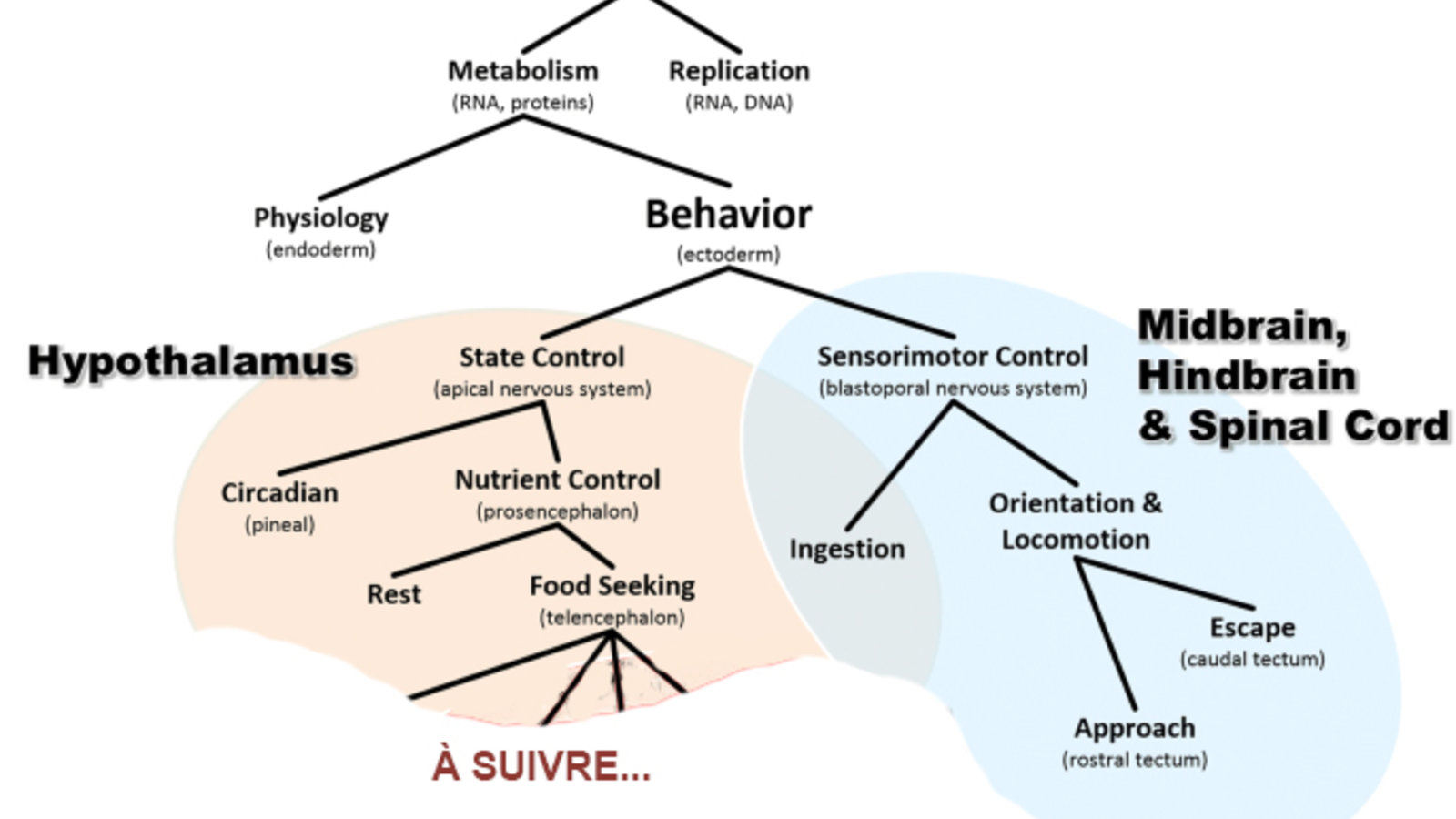
Tout en haut, on a évidemment l’apparition de la vie qu’on peut décliner en deux caractéristiques principales : d’un côté on a tout ce qui concerne le métabolisme, le maintien de la structure, l’autopoïèse quoi; et de l’autre on a la mémoire génétique de tout ça, l’ADN et tout ce qui touche à la reproduction (voir l’image en haut de ce billet). Ensuite on va distinguer encore deux choses dans le métabolisme : la physiologie, c’est-à-dire les régulations internes, des hormones entre autres, qui vont assurer l’équilibre de ce milieu intérieur; et les comportements qui sont ni plus ni moins que des boucles de régulation, mais à l’extérieur de l’organisme. Des boucles qui impliquent donc un mouvement, et donc un comportement, pour éventuellement se rapprocher d’une ressource et se l’approprier.
C’est cette branche des comportements que nous allons maintenant suivre pour parler concrètement des premiers animaux multicellulaires avec un système nerveux rudimentaire. Ils ressemblaient un peu au stade de la gastrula de notre développement embryonnaire. Chez ces animaux encore plus ou moins sphériques, comme les méduses ou les anémones de mer d’aujourd’hui, les neurones se répartissent déjà en deux grandes régions. D’abord la région apicale qui est riche en cellules sensibles aux molécules chimiques en solution et à la lumière. C’est elle qui va s’occuper des états comportementaux de base comme la gestion de l’énergie ou les cycles d’activité et de repos. Tout ça en sécrétant diverses neurohormones, étant donné que système nerveux et hormonal ne sont pas encore vraiment différenciés.
Et puis, à l’autre bout de l’animal, d’autres neurones vont contrôler les contractions rythmiques qui produisent soit l’aspiration d’eau dans l’organisme ou la propulsion par l’éjection rapide de cette eau. Ces deux débuts de spécialisation du système nerveux vont donner lieu à deux grandes familles de comportements. Dans le premier cas, un comportement qui va contrôler l’état général de l’organisme, ce qu’il a besoin pour demeurer dans un état viable pour son métabolisme. Et dans le deuxième cas, un type de comportement plus tourné vers l’environnement, vers ce qui s’y trouve comme ressource intéressante pour cet organisme. Autrement dit, une branche de nos comportements plus tournée vers l’exploitation des ressources autour de l’animal; et une autre tournée plus vers l’exploration de l’environnement pour trouver en premier lieu ces ressources et, inévitablement, éviter les dangers rencontrés.
Ce qui est particulièrement intéressant ici, c’est qu’on a pu montrer que chez plusieurs de ces animaux primitifs, c’est déjà la molécule de dopamine qui est sécrétée pour favoriser la recherche de nourriture et son exploitation dans l’environnement immédiat de l’animal. Donc un rôle très proche de celui que la dopamine joue encore dans la recherche de récompense chez les mammifères et les humains. Cela montre à quel point des choses apparues très tôt dans l’évolution peuvent être bien conservées jusqu’à nous
Je n’ai pas le temps d’entrer ici dans tous les détails de l’article, mais on arrive bientôt aux précurseurs des vertébrés. En gros, leur système nerveux va suivre la forme de leur corps allongé et leur cerveau va être constitué de structures nerveuses rudimentaires ressemblant à un hypothalamus attaché à un tronc cérébral et une moelle épinière qui vont assurer la nage ondulatoire de ces organismes. Le point important ici, c’est de constater qu’une des fonctions fondamentales du cerveau de ces animaux était de permettre l’alternance entre sa locomotion et son autre état principal de non locomotion, le comportement d’alimentation. Et cette opposition-là va demeurer présente jusqu’à nous.
Par la suite, avec l’avènement des premiers vertébrés, des poissons apparus il y a 530 millions d’années, la région apicale du système nerveux, va continuer d’être impliquée dans l’exploitation des ressources pour maintenir l’état d’équilibre intérieur de l’organisme et va devenir ce qu’on appelle aujourd’hui l’hypothalamus qui va aussi contrôler la température les cycles circadiens liés à la succession des jours et des nuits. Toujours chez les vertébrés, du côté de la locomotion assurée par le tronc cérébral et la moelle épinière va s’ajouter ce qui va bientôt devenir le mésencéphale. Et ça, ça va se faire en parallèle avec l’apparition de deux yeux, un de chaque côté de la tête, au lieu du spot unique de cellules photosensibles qu’il y avait avant chez les organismes plus primitifs. Quand ces yeux vont commencer à avoir une surface rétinienne capable de créer une carte topographique en deux dimensions de ce qui se passe dans le monde extérieur, la partie supérieure arrière du mésencéphale, le tectum caudal, va recevoir ces afférences et prendre de l’expansion. De sorte qu’il y aura bientôt dans ce tectum une carte capable d’orienter les réponses de fuite dans la direction opposée aux menaces. Et s’il y a deux menaces, le tectum va faire immédiatement une sorte de moyenne topographique qui va amener l’organisme à se faufiler entre les deux.
La partie avant ou rostrale du tectum va produire pour sa part les mouvements d’orientation vers les ressources d’intérêt pour l’animal. Autrement dit, elle va coordonner les mouvements d’approche, toujours en lien avec ce que les indices visuels de la rétine lui fournissent. Mais dans ce cas-ci, les neurones du tectum rostral ne vont pas fournir une moyenne comme output, ce qui serait complètement mésadapté dès qu’il y aurait plus qu’une source de nourriture. L’animal tendrait alors à se diriger entre les deux, et donc n’en attraperait aucune. Donc ici le calcul va plutôt être de type « winner-take-all », comme on dit en anglais.
Cette idée de compétition entre les neurones où un groupe particulier finit par gagner et remporter toute la mise, c’est-à-dire devenir la seule commande motrice effective, va devenir un principe très général pour la sélection des comportements. C’est le cas pour par exemple de la grenouille dont les circuits du tectum agissent comme un véritable détecteur pour les moustiques qui passent à proximité. Et ça peut se faire avec des mécanismes de base aussi simple que l’inhibition latérale, quand des neurones inhibent l’activité de leurs voisins, par exemple. On retrouve ça déjà dans le tectum d’animaux aussi anciens que la lamproie. Et ça peut produire ce type de réponse « winner-take-all » comme on va voir que c’est aussi le cas à plein d’endroits dans le cerveau humain.
Si vous me permettez une dernière observation qui m’a intéressée pour cette semaine, j’ajouterais qu’à partir des mammifères, le tectum optique va devenir ce qu’on appelle le collicule supérieur. Et quand, chez le rongeur, on stimule sa partie médiane, qui reçoit son information du champ visuel au-dessus de l’animal, ça produit des comportements défensifs et d’évitement. Mais quand on stimule la partie latérale du collicule supérieur, qui reçoit plus son information du bas du champ visuel de l’animal, ça produit des comportements appétitifs d’approche. La beauté de tout ça c’est que pour un rongeur, les prédateurs s’approchent pas mal toujours d’en haut et la nourriture se trouve pas mal tout le temps par terre, vers le bas.
Cela est tout à fait cohérent d’un point de vue évolutif et donne une idée de ce que des gens comme Karl Friston veulent dire quand ils parlent du corps d’un animal comme d’un modèle de son environnement. On poursuivra donc sur l’élaboration évolutive de ces modèles corps-cerveau à la base de nos comportement la semaine prochaine.

Vous aimerez aussi
-
 Rapport de l’ONU sur l’IA : un potentiel sous surveillanceMercredi 22 juillet 2026
Rapport de l’ONU sur l’IA : un potentiel sous surveillanceMercredi 22 juillet 2026 -
 Assis trop longtemps : risque accru de décès par cancerMercredi 22 juillet 2026
Assis trop longtemps : risque accru de décès par cancerMercredi 22 juillet 2026 -
 Un nouveau rôle pour les ovaires après la ménopauseLundi 20 juillet 2026
Un nouveau rôle pour les ovaires après la ménopauseLundi 20 juillet 2026 -
 Le réchauffement climatique nuit à la santé pulmonaireLundi 20 juillet 2026
Le réchauffement climatique nuit à la santé pulmonaireLundi 20 juillet 2026 -
 Diagrammes de Penrose-Carter et trous noirs en rotationDimanche 19 juillet 2026
Diagrammes de Penrose-Carter et trous noirs en rotationDimanche 19 juillet 2026 -
 Santé : quand l’IA perpétue les préjugésVendredi 17 juillet 2026
Santé : quand l’IA perpétue les préjugésVendredi 17 juillet 2026

Les plus populaires