Je poursuis cette semaine l’écriture du « journal de bord » du livre sur le cerveau et les comportements humains que j’ai écrit durant la pandémie. Commencé en janvier dernier dans la foulée du 20e anniversaire du Cerveau à tous les niveaux, ce journal me permet de vous faire part de mes réflexions au fil de la relecture du bouquin. Après le chapitre 1 qui portait sur le « connais-toi toi-même » de Socrate à l’heure des sciences cognitives et le chapitre 2 sur l’évolution cosmique, chimique et biologique qui a permis l’émergence des systèmes nerveux, je viens de terminer la réécriture du chapitre 3 sur notre grammaire neuronale. J’ai raconté maintes fois cette histoire déjà. C’est ma formation de base, l’électrophysiologie des mécanismes de la communication neuronale. J’en parle un peu plus loin dans le livre de « mon histoire d’amour avec mon mollusque », l’aplysie, dont j’avais caractérisé durant ma maîtrise un groupe de neurones sensoriels.
J’ai eu beau pendant deux ans de ma vie empaler quasi quotidiennement des neurones pour enregistrer leur activité électrique, je ne me suis jamais habitué à leur caractéristique sans doute la plus étonnante : leur capacité d’intégration de tous les signaux qui lui parviennent à tout moment afin de déterminer à chaque instant si ce neurone-là va, à son tour, émettre ou non un influx nerveux. J’ai coutume de résumer ça avec une citation que j’ai toujours trouvé très belle et que je traduis ici librement :
« Le fait qu’une cellule vivante se soit adaptée en une structure capable de recevoir et d’intégrer des données, de prendre des décisions fondées sur ces données, et d’envoyer des signaux aux autres cellules en fonction du résultat de cette intégration est un exploit remarquable de l’évolution. ». – Dharmendra Modha
Bien entendu le mot décision, qu’on associe spontanément à une décision humaine consciente, ne décrit pas ce qui se passe dans chacun de nos neurones. Parce que l’intégration qui s’y déroule (un terme qui décrit pas mal mieux ce qui se passe vraiment) va se faire tout seul. C’est-à-dire que les excitations et les inhibitions que reçoit le neurone à tout moment vont s’additionner et se soustraire comme on vient de le dire, mais ce qui va déterminer si un autre influx nerveux va repartir dans le second neurone, c’est le niveau du potentiel de membrane à un endroit très précis : le tout début de l’axone qui part du corps cellulaire. Cet à cet endroit que les canaux ioniques qui se trouvent dans la membrane vont changer pour devenir les canaux dépendants du voltage typiques de l’axone. Et donc si la somme des potentiels excitateurs et inhibiteurs en provenance de toutes les synapses du neurone est supérieure à cet endroit précis au seuil de déclenchement de l’influx nerveux, autour de – 55 à – 50 millivolts, eh bien un potentiel d’action va être généré.
Abonnez-vous à notre infolettre!
Pour ne rien rater de l'actualité scientifique et tout savoir sur nos efforts pour lutter contre les fausses nouvelles et la désinformation!
C’est pour ça qu’on appelle parfois cette région au début de l’axone la « zone gâchette », parce que c’est de là que part pour ainsi dire le coup de feu de l’influx nerveux. Et donc on voit comment la « décision », entre guillemets, d’émettre ou non un influx nerveux va dépendre à chaque instant de tout ce que le neurone est en train de recevoir en termes d’excitation ou d’inhibition partout sur ses dendrites et son corps cellulaire.
Et c’est comme ça qu’on en est venu à distinguer des phénomènes de base de l’intégration neuronale. Ce qu’on appelle par exemple la sommation temporelle, quand deux stimuli arrivent à une même synapse très vite l’un après l’autre. La dépolarisation causée par le premier ne s’est pas encore estompée quand le deuxième arrive, qui embarque en quelque sorte par-dessus, ce qui augmente les chances d’atteindre le seuil de déclenchement du potentiel d’action. Ou bien ce qu’on appelle la sommation spatiale, c’est-à-dire le fait qu’à un moment donné, il y a des excitations ou des inhibitions qui arrivent en même temps mais à différents endroits sur les dendrites ou le corps cellulaire, et que tout ça va s’additionner ou se soustraire au fur et à mesure de leur diffusion le long de la membrane. Et donc il faut voir le potentiel de membrane d’un neurone comme quelque chose de dynamique qui fluctue constamment au gré des excitations et des inhibitions qu’un neurone reçoit. Et produisant un potentiel d’action chaque fois que ce potentiel de membrane dépasse le seuil au niveau de la zone gâchette de l’axone.
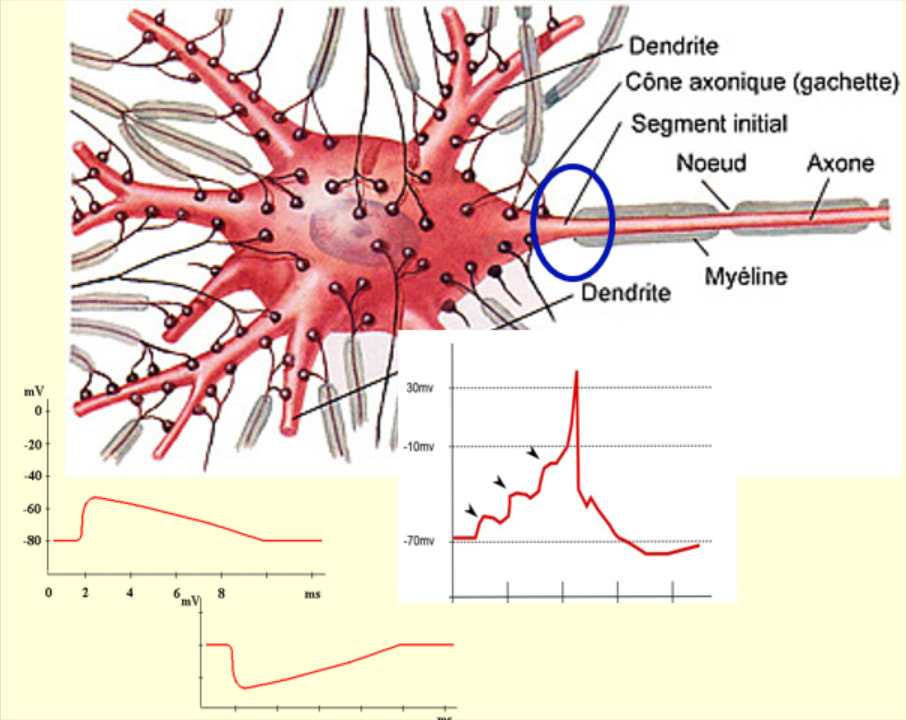
Cette intégration-là de tous ces signaux permet au neurone d’effectuer des opérations logiques de base comme ET, OU, NON-ET, etc. En fait, des travaux récents montrent que même de simples bouts de dendrites de certains neurones arrivent à faire ça !
Si on prend par exemple les deux exemples précédents de sommation temporelle ou spatiale qui réussissent à produire un influx nerveux. Ça peut être vu comme une fonction logique ET dans le sens où, dans une fenêtre temporelle donnée, ça prend l’activation de telle synapse ET de telle autre pour que le neurone réponde. On peut aussi qualifier ce type de computation de « détecteur de coïncidence ». On verra plus loin dans le livre que ça joue un rôle important dans certains types d’apprentissage.
De la même façon, si on a un neurone dont les influx nerveux sont déclenchés suite à l’activation de synapses sur tel dendrite OU tel autre, tu as ce qu’il faut pour implémenter la fonction logique OU. Et on peut en imaginer d’autres, qui combinent différents événements sur les dendrites. Par exemple si on a une synapse inhibitrice située sur la voie d’une excitation suffisante pour provoquer le déclenchement d’un influx nerveux, mais que cette inhibition est assez forte pour l’empêcher, on a ici comme un veto possible par cette connexion inhibitrice si elle est active. Ces capacités de calcul des neurones ont pas mal alimenté les analogies entre le cerveau est les ordinateurs, pour le meilleur, et surtout pour le pire.
Tout ça pour dire que la quantité d’influx nerveux que va émettre un neurone pendant un certain temps reflète ce qui s’est passé dans son réseau dendritique durant cette période. Et donc ce nombre de potentiel d’action par unité de temps, une seconde par exemple, est porteur d’information. Le taux de décharge d’un neurone, comme on l’appelle, est donc un des véhicules principaux dont disposent les neurones pour s’échanger de l’information. On a beaucoup mis l’accent là-dessus au début, mais on sait maintenant que les neurones peuvent s’échanger de l’information par d’autres stratégies, notamment en adoptant une activité rythmique et en synchronisant ces oscillations avec d’autres neurones, comme on va le voir aussi dans un chapitre ultérieur du livre.
Je vais arrêter ici pour l’avant-goût de ce 3e chapitre. Sinon pour ajouter qu’après avoir écrit pendant 20 ans dans un site web et dans un blogue, le retour à un support papier avec l’impossibilité de faire des hyperliens quand on réfère à d’autres chapitres ou à des articles en référence me frustre parfois un peu. Mais on a trouvé quelques trucs dans le livre pour référer simplement à des pages du site et du blogue du Cerveau à tous les niveaux, ainsi qu’aux nombreux articles en référence. Parce que je vois le livre, le site et le blogue comme formant un tout organique, intimement interreliés, où les limites de chacun sont compensés par les avantages des autres.

Vous aimerez aussi
-
 Dangereuses crèmes solaires sur TikTokVendredi 19 juin 2026
Dangereuses crèmes solaires sur TikTokVendredi 19 juin 2026 -
 Comment calculer le réchauffement supplémentaire causé par le réchauffementVendredi 19 juin 2026
Comment calculer le réchauffement supplémentaire causé par le réchauffementVendredi 19 juin 2026 -
 Avenir sombre pour les jeunes scientifiques aux États-UnisJeudi 18 juin 2026
Avenir sombre pour les jeunes scientifiques aux États-UnisJeudi 18 juin 2026 -
 A-t-on besoin de 2 jambes dans l’espace?Jeudi 18 juin 2026
A-t-on besoin de 2 jambes dans l’espace?Jeudi 18 juin 2026 -
 Une infrastructure souterraine vitale pour les écosystèmesMercredi 17 juin 2026
Une infrastructure souterraine vitale pour les écosystèmesMercredi 17 juin 2026 -


Les plus populaires